Anatomi tumbuhan-Anatomi tumbuhan adalah melihat keseluruhan fisik sebagai bagian-bagian yang secara
fungsional berbeda. Anatomi tumbuhan biasanya dibagi menjadi tiga bagian
berdasarkan hierarki dalam kehidupan:
- Organologi, mempelajari struktur dan fungsi organ berdasarkan jaringan-jaringan penyusunnya;
- Histologi, mempelajari struktur dan fungsi berbagai jaringan berdasarkan bentuk dan peran sel penyusunnya; dan
- Sitologi, mempelajari struktur dan fungsi sel serta organel-organel di dalamnya, proses kehidupan dalam sel, serta hubungan antara satu sel dengan sel yang lainnya. Sitologi dikenal juga sebagai biologi sel.
- A. AKAR

Secara umum, ada dua jenis akar yaitu:
- Akar serabut : Akar ini umumnya terdapat pada tumbuhan monokotil. Walaupun terkadang, tumbuhan dikotil juga memilikinya (dengan catatan, tumbuhan dikotil tersebut dikembangbiakkan dengan cara cangkok, atau stek). Fungsi utama akar serabut adalah untuk memperkokoh berdirinya tumbuhan.
- Akar tunggang : Akar ini umumnya terdapat pada tumbuhan dikotil. Fungsi utamanya adalah untuk menyimpan makanan.
Anatomi akar terdiri dari:
- Kambium : lapisan sel hidup pada tumbuhan yang aktif membelah berfungsi untuk memperbesar batang, terletak di antara kulit dan kayu.
- Pembuluh tapis (floem) : deretan sel yang dindingnya searah dengan poros akar – batang dan berlubang – lubang halus sehingga membentuk pembuluh. Fungsinya untuk mengangkut zat makanan dari akar keseluruh tubuh tumbuhan.
- Pembuluh kayu (xylem) : deretan sel yang dindingnya searah dengan poros akar – batang dan menyatu. Fungsinya untuk menyalurkan air yang mengandung mineral dari akar ke daun dan bagian lain tubuh.
B. BATANG
Batang tumbuhan mempunyai bentuk berbeda – beda. Pada umumnya batang dibagi menjadi tiga golongan, yaitu :
- Batang Berkayu
Tumbuhan jenis ini
biasanya besar, tinggi, dan bercabang – cabang. Daun tumbuhan ini
biasanya rimbun. Contohnya : mangga, jati, pohon jambu, dan lainnya.
- Batang Tidak Berkayu
Tumbuhan jenis ini biasanya tidak terlalu tinggi dan daunnya menempel pada batang. Contohnya : jagung dan tebu.
- Batang Semu
Tumbuhan jenis ini berupa pelepah – pelepah yang membentuk batang. Contohnya : pisang.
Keterangan struktur anatomi batang, yaitu :
- Epidermis : epidermis batang mempunyai sel – sel silika dan sel – sel gabus, misalnya pada batang tebu (Saccharum officinarum), dan kadang – kadang di lapisi oleh sel kutikula.
- Periderm : selaput luar epidermis yang terdapat di sekeliling mulut membentuk tonjolan berbentuk piala.
- Kortek : lapisan luar suatu organ, pada tumbuhan di bawah epidermis sebelah luar silinder pusat, terdiri dari sel – sel parenkim.
- Floem primer : dibentuk oleh prokambium ujung batang dan akar.
- Floem sekunder : terdiri dari unsur trakeal, serabut xylem dan parenkim kayu.
- Kambium : lapisan sel hidup terletak di kulit dan kayu, yang membuat jaringan kayu baru ke sebelah dalam dan membuat jaringan kulit baru ke sebelah luar. Fungsinya untuk memperbesar batang.
- Xylem sekunder : terdiri dari unsur trakeal, serabut xylem dan parenkim kayu.
- Xylem primer : dibentuk oleh prokambium ujung batang dan akar.
C. DAUN
Daun mempunyai bentuk yang bermacam – macam. Bentuk daun dapat dibedakan menjadi 5 macam, yaitu :
ø Bentuk bulat atau bundar : teratai besar.
ø Bentuk perisai : daun jarak.
ø Bentuk jorong : daun nangka dan nyamplung.
ø Bentuk memanjang : daun sirkaya dan sirsak.
ø Bentuk lanset : daun kamboja.
Keterangan gambar anatomi daun, yaitu :
- Epidermis terbagi atas epidermis atas dan epidermis bawah. Epidermis berfungsi melindungi jaringan di bawahnya.
- Jaringan palisade atau jaringan tiang adalah jaringan yang berfungsi sebagai tempat terjadinya fotosintesis
- Jaringan spons atau jaringan bunga karang yang berongga. Jaringan ini berfungsi sebagai tempat menyimpan cadangan makanan.
- Berkas pembuluh angkut yang terdiri dari xilem atau pembuluh kayu dan floem atau pembuluh tapis. Xilem berfungsi untuk mengangkut air dan garam-garaman yang diserap akar dari dalam tanah ke daun (untuk digunakan sebagai bahan fotosintesis). Sedangkan floem berfungsi untuk mengangkut hasil fotosintesis ke seluruh tubuh tumbuhan.
- Stoma (jamak: stomata) berfungsi sebagai organ respirasi. Stoma mengambil CO2 dari udara untuk dijadikan bahan fotosintesis. Kemudian stoma akan mengeluarkan O2 sebagai hasil fotosintesis. Stoma ibarat hidung kita dimana stoma mengambil CO2 dari udara dan mengeluarkan O2, sedangkan hidung mengambil O2 dan mengeluarkan CO2. Stoma terletak di epidermis bawah. Selain stoma, tumbuhan tingkat tinggi juga bernafas melalui lentisel yang terletak pada batang.
—D. BUNGA
Mahkota bunga : untuk memikat serangga yang menolong penyerbukan.
Benang sari : merupakan alat kelamin jantan pada tumbuhan, mengandung tepung sari.
Kelopak : pembungkus bunga selagi kuncup.
Putik : alat kelamin betina pada tumbuhan, yang akan menjadi bakal buah.
Dasar bunga : terletak di pangkal bunga, tempat melekatnya perhiasan bunga dan alat pembiakan.
Tangkai bunga : tempat melekatnya bunga.
—E. BUAH
Buah
merupakan perkembangan dinding bakal buah dan terkadang juga bagian –
bagian bunga lainnya. Buah terdiri atas kulit buah, daging buah dan
biji.
F. BIJI
Keterangan struktur anatomi biji, yaitu :
- Kulit biji : terletak di bagian luar biji dan melapisi seluruh bagian biji.
- Hipokotil : bagian bawah aksis (pangkal) yang melekat pada kotiledon.
- Radikula : bagian terminal (ujung).
- Epikotil : bagian atas pangkal.
- Plumula : bagian ujung, yaitu pucuk dengan sepasang daun.
Fisiologi tumbuhan adalah cabang botani yang mempelajari bekerjanya sistem kehidupan di dalam tubuh tumbuhan dan tanggapan terhadap pengaruh lingkungan sekitarnya. Seperti juga fisiologi hewan, fisiologi menggabungkan aspek fisika, kimiawi, dan biologi. Dari fisiologi tumbuhan ini lahirlah cabang-cabang campuran biologi, seperti biokimia dan biofisika. Fisiologi juga sangat mempengaruhi perkembangan genetika.
Objek kajian dalam fisiologi tumbuhan adalah fisika sel dan biofisika organ, fotosintesis, transportasi hara dan hasil metabolisme, regulasi pertumbuhan dan perkembangan, dan mekanisme respons terhadap rangsangan lingkungan.
Fisiologi tumbuhan diterapkan dalam pertanian untuk meningkatkan nilai produk hasil bumi. Beberapa contoh hasil kajian fisiologi yang diterapkan di pertanian adalah teknologi pemberian pupuk kimia untuk meningkatkan hasil dan penggunaan zat pengatur tumbuh untuk merangsang keserempakan pembungaan. Hasil penelitian di bidang fisiologi juga dapat dipakai untuk mendukung program pemuliaan tanaman, misalnya dalam merakit kultivar yang tahan kekeringan.
Pertumbuhan sel pada tumbuhan tingkat tinggi sering terjadi tanpa
peningkatan volume sitosol. Karena kekuatan ionik rendah dari dinding
sel, air cenderung untuk meninggalkannya dan memasuki sitosol dan
vakuola, menyebabkan sel untuk berkembang. Proses melonggarkan dari
dinding sel primer yang disebabkan oleh auksin, memungkinkan sel untuk
berkembang dalam arah tertentu, ukuran dan bentuk tumbuhan ditentukan
terutama oleh jumlah dan arah pembesaran ini (Gambar 1a). Sel-sel
individu tumbuhan dapat meningkatkan ukuran dengan sangat cepat dengan
melonggarkan dinding sel dan mendorong sitosol dan membran plasma.
Peningkatan volume sel hanya disebabkan perluasan vakuola intraselular
oleh serapan air. Kita dapat membuktikan fenomena ini dengan
mempertimbangkan bahwa jika semua sel di pohon redwood dikurangi seukuran sel hati (≈ 20 mm diameter), pohon itu hanya akan memiliki ketinggian maksimum 1 meter.
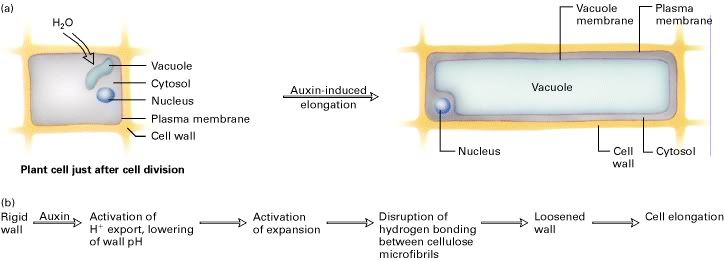
Gambar 1: Perpanjangan sel tumbuhan:
(a) Perubahan struktur sel tanaman selama proses perpanjangan. Serapan air menyebabkan tekanan internal (turgor), kehadiran auksin, dinding sel melonggar, dan tekanan turgor dinding pada dinding longga menyebabkan pemanjangan sel. (b) Usulan mekanisme dinding sel mengendur dalam sel tumbuhan. [Part (b) adapted from L. Taiz, 1994, Proc. Nat’l. Acad. Sci. USA 91:7387.]
Sumber: Section 22.5, The Dynamic Plant Cell Wall(a) Perubahan struktur sel tanaman selama proses perpanjangan. Serapan air menyebabkan tekanan internal (turgor), kehadiran auksin, dinding sel melonggar, dan tekanan turgor dinding pada dinding longga menyebabkan pemanjangan sel. (b) Usulan mekanisme dinding sel mengendur dalam sel tumbuhan. [Part (b) adapted from L. Taiz, 1994, Proc. Nat’l. Acad. Sci. USA 91:7387.]
Kemampuan auksin (indole-3-asam asetat) untuk memercepat induksi
perpanjangan sel pertama kali ditunjukkan dalam eksperimen klasik pad
coleoptile rumput dan gandum. Menurut hipotesis pertumbuhan-asam, auksin
merangsang sekresi proton pada akhir “pertumbuhan” pada sel dengan
mengaktifkan (langsung atau tidak langsung) suatu pompa proton yang
terikat pada membran plasma (Gambar1b). Akibatnya, pH dinding sel
sekitar daerah membran plasma jatuh dari 7,0 normal menjadi turun 4,5.
PH rendah mengaktifkan kelas protein
dinding sel, disebut expansin yang dapat mengganggu ikatan hidrogen
antara mikrofibril selulosa sehingga menyebabkan struktur lapisan dari
dinding sel untuk melonggar. Dengan kekakuan dinding berkurang, sel
dapat memanjang.
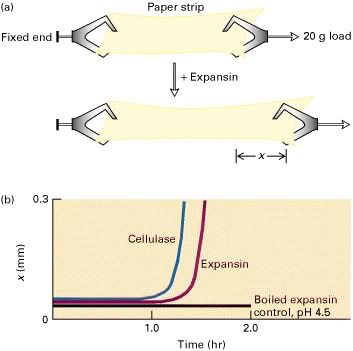
Gambar 2: Demonstrasi experimental diamana expansin mengendurkan ikatan hidrogen:
(a) Didalam elastometer, sebuah strip kertas dijepit pada kedua
ujungnya dan direndam dalam larutan. Salah satu ujung terpasang ke
beban, sementara ujung lainnya dipegang tetap. Suatu agen yang dapat
mengendurkan ikatan kovalen atau hidrogen antara serat selulosa akan
menyebabkan strip kertas memanjang sebanyak x kali. Gerakan penjepit
dicatat. (b) Perlakuan strip kertas dengan expansin pada pH 4,5 (merah)
mengakibatkan melemahnya dari molekul selulosa reversibel. Sebaliknya,
secara ireversibel selulase melemahkan kertas dengan memecah ikatan
kovalen dalam polimer. Percobaan kontrol menunjukkan bahwa pelemahan
tidak disebabkan oleh larutan dengan pH 4,5 dan itu tergantung pada
keaktifan protein.
Sumber:Section 22.5, The Dynamic Plant Cell Wall
Expansin ditemukan dan dimurnikan menggunakan uji biokimia baru pada
kertas selulosa murni. Kenapa kertas?, seperti dinding sel tanaman dari
mana itu dibuat sehingga memilki kekuatan mekanik antara ikatan hidrogen
dengan mikrofibril selulosa. Ekstrak dari dinding sel tumbuhan diuji
untuk kemampuanya untuk mekanisme melemahkan kertas pada nilai pH antara
3,0 dan 5,0, tetapi tidak pada pH 7 (Gambar 2). Expansin memicu
melonggarnya dinding sel dan berkebalikan ketika pH dinaikkan kembali ke
7,0. Hal ini menunjukkan expansin yang tidak memutuskan ikatan kovalen
dalam selulosa. Bukti tambahan untuk hipotesis pertumbuhan-asam berasal
dari studi tentang fusicoccin suatu senyawa jamur. Seperti auksin,
fusicoccin menginduksi pemanjangan sel yang cepat dan memicu memompa
proton keluar dari sel yang sensitif dengan disertai melonggarkan
dinding sel sekitar. Aktivitas fusicoccin atau auksin dapat diblokir
melalui mensisipi dinding sel dengan buffer yang mencegah pH
ekstraselular turun.
IAA merupakan hormon endogenous yang terdapat pada tanaman terbentuk dari triptofan dan terdapat cicin indol (Davies, 1995). Keunikan dari IAA adalah cicin tidak jenuh (rangkap dua) dan rantai samping asam. Terbentuk ruang yang tepat antara cicin dan rantai sampingnya (Gardner et al,. 1991). IAA memiliki peran fisiologis tumbuhan dalam pemanjangan dan perkembangan sel (Campbell dan Reece, 2002).
Auksin menyebabkan pembentangan jaringan korteks, floem dan kambium sehingga sel sklerenkim menjadi rusak memicu akar keluar. Pengaruh auksin pada perkembangan sel-sel pada daerah maristem menyebabkan sel-sel memanjang (Dwijoseputro, 1994).
Kisaran pengaruh auksin meliputi metabolisme selular sampai ke koordinasi morfogenesis tumbuhan termasuk absisi dan penuaan. Pengaruh selular seperti (Gardner et al,. 1991):
1. Peningkatan sintesis nukleotida DNA dan RNA
2. Sintesis protein dan enzim
3. Pertukaran proton
4. Potensial membran
5. Pengambilan kalium
Peran utama auksin adalah pemompaan proton pada membran plasma. Pada daerah perpanjangan sel, auksin meningkatkan pompa proton sehingga dalam beberapa menit meningkatkan potensial membran dan pH dalam didinding menurun (Gambar 2). Lingkungan yang menjadi asam menyebabkan aktivasi enzim ekspansin yang melepaskan ikatan hidrogen antara ikatan mikrofibril selulosa dan melonggarkan struktur dinding sel. Intergritas selulsa murni menjadi lemah akibat pengaruh auksin. Peningkatan potensial membran mengakibatkan pengambilan ion dari luar sel ke dalam sehingga terjadi pemasukan air secara osmosis. Pemasukkan air diiringi dengan plastisitas dinding sel mengakibatkan pemanjangan sel. Auksin menginduksi ekspresi gen untuk menghasilkan protein-protein baru pada daerah perpanjangan sel hanya dalam beberpa menit. Bebarapa protein bersifat mendorong atau menghambat transkripsi ekspresi gen lainnya. Tahap selanjutnya, sel akan membentuk sitoplasma dan bahan didind sel. Auksin juga merespon pada pertumbuhan lainnya (Campbell dan Reece, 2002).
IAA merupakan hormon endogenous yang terdapat pada tanaman terbentuk dari triptofan dan terdapat cicin indol (Davies, 1995). Keunikan dari IAA adalah cicin tidak jenuh (rangkap dua) dan rantai samping asam. Terbentuk ruang yang tepat antara cicin dan rantai sampingnya (Gardner et al,. 1991). IAA memiliki peran fisiologis tumbuhan dalam pemanjangan dan perkembangan sel (Campbell dan Reece, 2002).
Auksin menyebabkan pembentangan jaringan korteks, floem dan kambium sehingga sel sklerenkim menjadi rusak memicu akar keluar. Pengaruh auksin pada perkembangan sel-sel pada daerah maristem menyebabkan sel-sel memanjang (Dwijoseputro, 1994).
Kisaran pengaruh auksin meliputi metabolisme selular sampai ke koordinasi morfogenesis tumbuhan termasuk absisi dan penuaan. Pengaruh selular seperti (Gardner et al,. 1991):
1. Peningkatan sintesis nukleotida DNA dan RNA
2. Sintesis protein dan enzim
3. Pertukaran proton
4. Potensial membran
5. Pengambilan kalium
Peran utama auksin adalah pemompaan proton pada membran plasma. Pada daerah perpanjangan sel, auksin meningkatkan pompa proton sehingga dalam beberapa menit meningkatkan potensial membran dan pH dalam didinding menurun (Gambar 2). Lingkungan yang menjadi asam menyebabkan aktivasi enzim ekspansin yang melepaskan ikatan hidrogen antara ikatan mikrofibril selulosa dan melonggarkan struktur dinding sel. Intergritas selulsa murni menjadi lemah akibat pengaruh auksin. Peningkatan potensial membran mengakibatkan pengambilan ion dari luar sel ke dalam sehingga terjadi pemasukan air secara osmosis. Pemasukkan air diiringi dengan plastisitas dinding sel mengakibatkan pemanjangan sel. Auksin menginduksi ekspresi gen untuk menghasilkan protein-protein baru pada daerah perpanjangan sel hanya dalam beberpa menit. Bebarapa protein bersifat mendorong atau menghambat transkripsi ekspresi gen lainnya. Tahap selanjutnya, sel akan membentuk sitoplasma dan bahan didind sel. Auksin juga merespon pada pertumbuhan lainnya (Campbell dan Reece, 2002).

Gambar: Perpanjangan sel merupakan respon terhadap auksin. Suatu hipotesis pertumbuhan asam (Acid Growth Hipotesis) (Campbell dan Recce, 2002).
Sumber:glogster.com
Terdapat dua mekanisme sintesis IAA:
1. Eliminasi asam amino
2. Eliminasi ujung gugus karboksil dari cicn samping triptofan
Mekanisme yang paling diketahui adalah:
a. Reaksi transaminasi : gugus amino bergabung dengan α-keto menjadi asam indol
asetat
b. Dekarboksilasi : indolpiruvat menjadi indolasetaldehida.
c. Oksidasi : indolasetaldehid diubah menjadi IAA
Enzim yang diperlukan untuk mengubah triptofan menjadi IAA terdapat pada jaringan muda, seperti maritem ujung, daun muda, dan buah dalam pertumbuhan.
1. Eliminasi asam amino
2. Eliminasi ujung gugus karboksil dari cicn samping triptofan
Mekanisme yang paling diketahui adalah:
a. Reaksi transaminasi : gugus amino bergabung dengan α-keto menjadi asam indol
asetat
b. Dekarboksilasi : indolpiruvat menjadi indolasetaldehida.
c. Oksidasi : indolasetaldehid diubah menjadi IAA
Enzim yang diperlukan untuk mengubah triptofan menjadi IAA terdapat pada jaringan muda, seperti maritem ujung, daun muda, dan buah dalam pertumbuhan.

Gambar: skema jalur bioseintesis IAA (Salisbury dan Ross, 1992)
Zat pengatur tumbuh (ZPT) seperti asam geberelin dan IAA memilki efek
fisiologis terhadap pemecahan dormansi biji memelalui mekanisme
perkecambahan. Menurut Salisbury dan Ross (1995)gormon tersebut akan
masuk kedalam sel dan akan ditangkap oleh protein reseptor .
Gambar: Mekanisme hormon ZPT masuk kedalam sel
Sumber: bio.miami.eduProtein reseptor akan mengaktifkan enzim fosfolipase c (PLC). PLC menghidrolisis lipid yang mengandung inositol (fosfoinositida). Selain itu, PLC juga menghidrolisis jenis lipid terakhir berupa fosfatdilinositol 4,5-bifosfat (PIP2) mengahsilkan inositol-1,3,5-trifosfat (IP3) dan diasilgliserol (DAG). IP3 mengiduksi lepasnya Ca2+ ke sitosol yang tersimpan dalam vakuola. DAG berfungsi dalam membran sel dengan mengaktifkan enzimprotein kinase c (PKC). PKC memanfaatkan ATP untuk fosforilasi enzim tertentu untuk proses metabolisme. Sealain itu, peningkatan Ca2+ mengakibatkan aktivasi enzim lain termasuk PKC. Empat ion Ca2+ bergabung membentuk Ca-kalmodulin yang dapat menginduksi enzim kinase NAD+ dengan memfosforilasi menjadi NADP+ dan ATPase.
Produksi enzim disebabkan pengaruh hormon terhadap mRNA untuk mentranslasi enzim-enzim yang bersifat katalisis tinggi. Hormon dapat menyebabkan ekspresi gen dengan meningktkkan:
1. Aktivitas RNA polimerase
2. Jumlah RNA ribosom
3. Jumlah mRNA
Reseptor pada membran palasma akan mentranslokasi hormon menuju nuleus. Auksin menginduksi DNA dalam nukleoplasma untuk bertanskripsi menjadi pre-RNA (primary transcript) proses ini dikatalisis oleh enzim RNA polimerase. Melalui mekanisme RNA processing. Pre-RNA dirubah menjadi, mRNA, tRNA, rRNA,snRNA. snRNA(small nucleat RNA) membantu transport mRNA dari nuleoplasma menuju sitoplasma. Di dalam sitosol, mRNA terikat dalm ribosom dan memulai proses translasi protein. mRNA oleh ribosom akan diterjemahakan menjadi brbagai jenis enzim dan protein
AUKSIN
Auksin

Gambar: Struktur IAA
Sumber:www.plant-hormones.info
Alamiah auksin
Istilah auksin (gambar 1) berasal dari bahasa latin dari kata
“auxein” yang artinya tumbuh. Senyawa yang pada umumnya dianggap auksin
jika memiliki karakter kemampuan dalam menginduksi perpanjangan sel pada batang dan tersusun dari asam indolasetat
(auksin pertama yang diisolasi) dalam aktivitas fisiologisnya. Auksin
biasanya dapat mempengaruhi proses lainnya selain perpanjangan sel. Oleh
karena itu dianggap semua auksin berperanan kritis dan membantu
mendifinisikan hormon (Arteca, 1996; Mauseth, 1991; Raven, 1992;
Salisbury and Ross, 1992).
Sejarah auksin dan penelitian terdahulu
Auksin merupakan hormon tanaman yang pertama kali ditemukan. Charles
Darwin merupakan diantara ilmuwan pertama yang meneliti tentang hormon
tanaman. Pada bukunya “Kekuatan gerak tanaman” yang ditampilkan pada
tahun 1880. Dia merupakan orang yang pertama mendiskripsikan pengaruh
cahaya pada pergerakan coleoptile rumput kanari (Phalaris canariesnsis).
Koleoptil daun terspesialisasi yang berasal dari nodus pertama dimana
merupakan pembungkus epikotil tanaman pada tahap perkecambahan untuk
proteksi sampai muncul dari tanah. Ketika cahaya diarahkan pada
koleoptil, maka akan membungkuk sesuai dengan arah datangnya cahaya.
Ketika ujung koleoptil ditutupi dengan alumunium foil, maka tidak akan
terjadi pembungkukan menuju arah cahaya. Namun, jika ujung koleoptil
dibiarkan terbuka tetapi sebagian tepat dibawah ujung tetap tertutup,
maka paparan dari cahaya menghasilkan kelengkungan tanaman menuju
cahaya. Eksperimen Darwin ini membuktikan bahwa ujung koleoptil terdapat
jaringan yang bertanggung jawab untuk menerima cahaya dan memproduksi
beberapa signal diamana akan dikirimkan pada bagian bawahnya koleoptil
dimana respon fisiologis pembungkukan terjadi. Dia kemudian memotong
ujung dari koleoptil dan menyinari sisa koleoptil dengan cahaya untuk
melihat terjadinya proses pelengkungan. Kelengkungan tanaman tidak
terjadi sehingga melaporkan hasil dari eksperimen pertama Darwin ini
(Darwin, 1880).
Pada tahun 1885, Salkowski menemukan Indole-3-acetic acid (IAA) pada media fermentasi (Salkowski, 1885). Isolasi produk serupa dari jaringan tanaman tidak dapat ditemukan pada jaringan tanamn hampir selama 50 tahun. IAA merupakan auksin utama yang terlibat dalam banyak proses fisiologis pada tanaman (Arteca, 1996). Pada tahun 1907, kajian Fitting tentang pengaruh sayatan pada sisi terkena cahaya dan tidak terkena cahay pada tanaman. Hasilnya telah membantu dalam memahami jika proses translokasi signla benar-benar terjadi pada sisi khusus tanaman tetapi hasil ini kurang menyakinkan karena signal ini mampu melintasi atau menhilang dari sekitar sayatan ( Fitting, 1907). Pada tahun 1913, Boysen-jensen memodifikasi eksperimen Fitting melalui penyisipan potongan mika untuk memblok jalannya signal dan menujukan bahwa transport auksin menuju dasar telah terjadi pada sisi gelap tanaman berkebalikan dengan dengan sisi yang terekspos cahaya searah (Boysen-Jensen, 1913). Pada tahun 1918, Paal menegaskan hasil Boysen-Jansen melalui pemotongan ujung koleoptil pada kondisi gelap, menyinari hanya bagian ujung dengan cahaya, menggantikan ujung koleoptil pada tanaman tetapi bagian pangkal dipusatkan ke salah satu sisi atau lainnya. Hasilnya menunjukkan, bagian mana saja pada koleoptil yang terpapar cahaya, kelengkungan terjadi menuju pada sisi lainnya (Paal, 1918). Soding merupakan ilmuwan berikutnya yang memeusatkan perhatian terhadap penelitian auksin berdasarkan ide dari Paal. Dia menunjukan jika ujung dari koleoptil dipotong akan menghasilkan reduksi dari pertumbuhan tetapi jika dipotong dan kemudian pertumbuhan tetap berlanjut berganti pada tempat lainnya (Soding, 1925).
Pada tahun 1885, Salkowski menemukan Indole-3-acetic acid (IAA) pada media fermentasi (Salkowski, 1885). Isolasi produk serupa dari jaringan tanaman tidak dapat ditemukan pada jaringan tanamn hampir selama 50 tahun. IAA merupakan auksin utama yang terlibat dalam banyak proses fisiologis pada tanaman (Arteca, 1996). Pada tahun 1907, kajian Fitting tentang pengaruh sayatan pada sisi terkena cahaya dan tidak terkena cahay pada tanaman. Hasilnya telah membantu dalam memahami jika proses translokasi signla benar-benar terjadi pada sisi khusus tanaman tetapi hasil ini kurang menyakinkan karena signal ini mampu melintasi atau menhilang dari sekitar sayatan ( Fitting, 1907). Pada tahun 1913, Boysen-jensen memodifikasi eksperimen Fitting melalui penyisipan potongan mika untuk memblok jalannya signal dan menujukan bahwa transport auksin menuju dasar telah terjadi pada sisi gelap tanaman berkebalikan dengan dengan sisi yang terekspos cahaya searah (Boysen-Jensen, 1913). Pada tahun 1918, Paal menegaskan hasil Boysen-Jansen melalui pemotongan ujung koleoptil pada kondisi gelap, menyinari hanya bagian ujung dengan cahaya, menggantikan ujung koleoptil pada tanaman tetapi bagian pangkal dipusatkan ke salah satu sisi atau lainnya. Hasilnya menunjukkan, bagian mana saja pada koleoptil yang terpapar cahaya, kelengkungan terjadi menuju pada sisi lainnya (Paal, 1918). Soding merupakan ilmuwan berikutnya yang memeusatkan perhatian terhadap penelitian auksin berdasarkan ide dari Paal. Dia menunjukan jika ujung dari koleoptil dipotong akan menghasilkan reduksi dari pertumbuhan tetapi jika dipotong dan kemudian pertumbuhan tetap berlanjut berganti pada tempat lainnya (Soding, 1925).

Gambar: Percobaan Went
Sumber: http://www.plant-hormones.info
Pada tahun 1926, seorang mahasiwa pascasarjana dari Belanda yang
bernama Fritz Went mempublikasikan laporannya yang mendiskripsikan
bagaimana dia mengisolasi substansi pertumbuhan tanaman melalui cara
menempatkan agar blok dibawah ujung koleoptil selama waktu tertentu
kemudian dilepaskan dan menempatkannya pada batang Avena yang dipotong
(Went, 1926). Setelah penempatan agar tersebut, batang itu memulai
pertumbuhan kembali (Lihat Gambar 2). Pada tahun 1928, Went
mengembangkan metode untuk perhitungan substansi pertumbuhan tanaman.
Hasilnya menyatakan bahwa kelengkungan pada batang proposional dengan
jumlah substansi pertumbuhan pada agar (Went, 1928). Uji ini disebut tes
kelengkungan avena ( Lihat Gambar 3).

Gambar: Percobaan Went
Sumber: http://www.plant-hormones.info
Sebagian besar pengetahuan tentang auksin saat ini diperoleh dari
aplikasi ini. Pekerjaan Went memberikan pengaruh besar terhadap
penelitian tentang substansi pertumbuhan tanaman. Dia sering sikaitakn
dengan istilah auksin tetapi pada kenyataannya Kogl dan Haagen-smitlah
yang pertama memurnikan senyawa auxientriolic acid (auxin A) dari urin
manusia pada tahun 1931 (Kogl dan Haagen-Smit, 1931). Sealnjutnya Kogl
mengisolasi senyawa lainnya dari urin dimana mirip dengan struktur dan
fungsi auksin A, salah satunnya adalah Indole-3 acetic acid (IAA) yang
pada mulanya ditemukan oleh Salkowski pada tahun 1985. Pada tahun 1954,
komite ahli fisiologi tanaman merangkai karakter kelompok dari auksin.
Istilah auksin berasal dari bahasa yunani yang berarti “to grow”.
Senyawa yang dapat diaanggap sebagai auksin jika disintesis oleh tanaman
dan substansi dimana memilki aktivitas serupa dengan IAA (uksin pertama
yang diisolasi dari tanaman) (Arteca, 1996; Davies, 1995).
Biosintesis dan metabolisme auksin
IAA secara kemikal mirip dengan asam amino triptofan diamana dianggap
sebagai asal dari bentuk molekul IAA. Tiga mekanisme yang dapat
menjelaskan perubahan ini:
Enzim yang berperan dalam biosintesis IAA merupakan yang paling aktif pada jaringan muda seperti pada ujung maristem apikal dan daun yang sedang tumbuh serta buah. Jaringan serupa dimana ditemukan konsentrasi tinggi IAA. Salah cara tumbuhan mengontrol jumlah IAA yang ada pada jaringan pada waktu tertentu adalah melalui pengontrolan hormon biosintesis. Mekanisme kontrol lainnya melibatkan produksi secara konjugasi dimana molekul sederhana dirangkai menjadi hormon tetapi belum aktif. Pembentukkan konjugasi mungkin merupakan mekanisme penyimpanan dan penyaluran hormon hormon aktif. Konjugasi dapat dibentuk dari IAA melalui enzim hidrolase. Konjugasi dapat cepat teraktivasi ketika signal stimulus lingkungan mempercepat respon secara hormonal. Degradasi auksin merupakan akhir dari metode pengontrolan level auksin. Dua mekanisme proses ini seperti dibawah:
Oksidasi IAA oleh oksigen menghasilkan hilangnya kelompok karboksil dan 3-metilenoksindole sebagai produk utama pemecahan. IAA oksidase merupakan enzim dimana mengkatalisis proses ini. Konjugasi IAA dan auksin sintetik seperti 2,4-D tidak dapat dihancurkan melalui proses ini. C-2 dari cicin heterosiklik dioksidasi menghasilkan oxindole-3-acetic acid. C-3 kemudia dioksidasi dengan penambahan C-2 sehingga menghasilkan Dioxindole-3 acetic acid.
Mekanisme biosintesis dan degradasi dari molekul auksin sangat penting untuk aplikasi agrikultur dimasa depan. Informasi mengenai metabolisme auksin memicu manipulasi genetik dan kemia dari level hormon endogen sehingga menghasilkan pertumbuhan yang diinginkan dan deferensiasi pada spesies tumbuhan penting. Pada akhirnya, ada kemungkina untuk meregulasi pertumbuhan tumbuhan tanpa penggunaan herbisida dan pupuk yang berbahaya (Davies, 1995; Salisbury and Ross, 1992).
- Triptofan diubah menjadi asam indolpiruvat melalui reaksi transmisi.
- Asam indolepiruvat kemudian diubah menjadi indoleasetaldehid melalui reaksi dekarboksilasi.
- Tahap akhir merupakan oksidasi indoleasetaldehid menghasilkan asam indoleasetat.
Enzim yang berperan dalam biosintesis IAA merupakan yang paling aktif pada jaringan muda seperti pada ujung maristem apikal dan daun yang sedang tumbuh serta buah. Jaringan serupa dimana ditemukan konsentrasi tinggi IAA. Salah cara tumbuhan mengontrol jumlah IAA yang ada pada jaringan pada waktu tertentu adalah melalui pengontrolan hormon biosintesis. Mekanisme kontrol lainnya melibatkan produksi secara konjugasi dimana molekul sederhana dirangkai menjadi hormon tetapi belum aktif. Pembentukkan konjugasi mungkin merupakan mekanisme penyimpanan dan penyaluran hormon hormon aktif. Konjugasi dapat dibentuk dari IAA melalui enzim hidrolase. Konjugasi dapat cepat teraktivasi ketika signal stimulus lingkungan mempercepat respon secara hormonal. Degradasi auksin merupakan akhir dari metode pengontrolan level auksin. Dua mekanisme proses ini seperti dibawah:
Oksidasi IAA oleh oksigen menghasilkan hilangnya kelompok karboksil dan 3-metilenoksindole sebagai produk utama pemecahan. IAA oksidase merupakan enzim dimana mengkatalisis proses ini. Konjugasi IAA dan auksin sintetik seperti 2,4-D tidak dapat dihancurkan melalui proses ini. C-2 dari cicin heterosiklik dioksidasi menghasilkan oxindole-3-acetic acid. C-3 kemudia dioksidasi dengan penambahan C-2 sehingga menghasilkan Dioxindole-3 acetic acid.
Mekanisme biosintesis dan degradasi dari molekul auksin sangat penting untuk aplikasi agrikultur dimasa depan. Informasi mengenai metabolisme auksin memicu manipulasi genetik dan kemia dari level hormon endogen sehingga menghasilkan pertumbuhan yang diinginkan dan deferensiasi pada spesies tumbuhan penting. Pada akhirnya, ada kemungkina untuk meregulasi pertumbuhan tumbuhan tanpa penggunaan herbisida dan pupuk yang berbahaya (Davies, 1995; Salisbury and Ross, 1992).
Fungsi Auksin
Berikut ini merupakan beberapa respon auksin yang diketahui (Davies,
1995; Mauseth, 1991; Raven, 1992; Salisbury and Ross, 1992).1. Menstimulasi pemanjangan sel
2. Menstimulasi pembelahan sel pada kambium dan kombinasi dengan sitokinin pada jaringan
3. Menstimulasi deferensiasi floem dan xilem
4. Menstimulasi inisiasi akar pada pemtongan batang dan perkembangan akar lateral pada kultur jaringan
5. Memediasi respon tropistik pembungkukkan dalam menanggapi garvitasi dan cahaya
6. Auksin yang disuplai dari tunas apikal menekan pertumbuhan tunas lateral
7. Menunda penguguran daun
8. Dapat menghambat atau mempromosi pengguguran (melalui stimulasi etilen) daun dan buah
9. Dapat menginduksi pengaturan perbuahan dan pertumbuhan pada beberapa tanaman
10. Melibatkan pergerakkan asimilasi auksin akibat pengaruh transport floem
11. Menunda kematangan buah
12. Memicu perbungggan pad Bromeliad
13. Menstimulasi pertumbuhan pada bagian bunga
14. Memicu pengurangan sifat betina pada bunga deoecidous
15. Menstimulasi produksi etilen pada konsentrasi tinggi
Diatas telah menggambarkan pengaruh auksin pada perkembangan strawberry. Achenes telah memproduksi auksin. Ketika dilepaskan, strawberry tidak dapat berkembang (Raven, 1992).
0 Response to "Anatomi dan Fisiologi Tumbuhan"
Post a Comment